Mise à jour: VT suit les statistiques COVID et la désinformation / censure depuis le premier jour. Maintenant,
à la lumière de l'attaque de Beyrouth par Israël (Trump nous soutient sur ce fait), nous
considérons maintenant la sortie d'une version hybridée de COVID 19 sur
la zone métropolitaine de New York comme une attaque terroriste.
Les
statistiques sur les hospitalisations et les décès ont été modifiées /
censurées, ce qui prouve qu'il s'agissait d'une attaque biologique. Les
taux de mortalité étaient plusieurs fois plus élevés qu'ailleurs et
ciblaient même spécifiquement la population juive, ce qu'Israël a déjà
fait.
Nous
avons observé la manipulation d'Internet par Google Corporation, nous
les avons suivis et avons trouvé des preuves claires et absolues qui
relient ceux qui ont exécuté l'attaque de Beyrouth à ceux qui ont diffusé
COVID 19 aux États-Unis et dans le monde.
Nous
avons également regardé des vidéos de Beyrouth censurées sur Facebook,
Google / YouTube et Twitter et nous nous sommes coordonnés avec MSM pour
vendre un faux récit impliquant un engrais non explosible alors que les
preuves d'une attaque massive non seulement d'Israël, mais aussi d'un
brillant ensemble de drones, d'avions et même un missile.
Qui sait, une bombe nucléaire aurait pu y être chargée facilement dès la sortie d'un camion et personne ne le remarquerait.
Maintenant
que Beyrouth est en ruine, que les Émirats arabes unis rejoignent le
Grand Israël et que la monstruosité de Qanon vend un État policier à la
populace, l'inexplicable devient clair.
Ci-dessous,
nous prouvons la création de COVID, un projet de la CIA réalisé à
travers un programme de couverture dans une université privée utilisant
des experts en armes biologiques. Maintenant, on le voit sorti, la cible? Piller
l'économie américaine, amener l'Amérique à la guerre civile et
permettre au pouvoir de se centraliser sous le contrôle de la Casher Nostra
(alias Mafia juive, alias Yiddish Connetion) sans subterfuge inutile.
Cet
article contient des preuves tangibles qui ne peuvent être ni
contestées ni refusées, que vous pouvez soumettre à toute agence
gouvernementale ou professionnel de la santé.
Ce
qui n'est pas encore prouvé, mais qui vient à l'esprit, c'est que le
programme américain d'armes biologiques à Fort Detrick, Maryland,
l'équipement et certainement le personnel clé, ont certainement migré
vers les laboratoires secrets des grandes universités d'État afin de «se
cacher à la vue de tous».
Suivez les carrières, tous les liens sont inclus, de ceux qui ont travaillé sur le projet Wuhan-COVID en 2017.
De
plus, notez que le même personnel et le même équipement sont utilisés
pour de fausses recherches et tests de «prévention» que la
militarisation et la production réelle.
Depuis
la rédaction de cet article, nous avons commencé à examiner les
opérations mondiales de l'entrepreneur nucléaire / bio / chimie
américain, le favori de Kushner-Trump, Battelle, et leurs laboratoires
secrets à travers le monde.
Quand nous avons commencé, notre peuple a commencé à être menacé. C'était une grave erreur.
Soumettez ce document à tout médecin ou à tout autre spécialiste qualifié en bio-sciences. Voyez ce qu'ils disent.

POUR PARTAGER CET ARTICLE SUR FACEBOOK, VEUILLEZ COPIER et COLLER ce lien: https://www.veteranstodaynetwork.com/2020/04/29/documentary-proof-university-of-north-carolina-generated-covid-19/
introduction
Les
documents ci-dessous montreront que la recherche pour créer COVID 19 a
commencé aux États-Unis en 2006 et a abouti à une arme biologique
réussie en 2015, avec des travaux effectués à l'Université de Caroline
du Nord et à Harvard et au laboratoire de la Food and Drug
Administration en Arkansas. .
Leur travail était intitulé:
Un groupe de coronavirus de chauve-souris en circulation de type SRAS montre un potentiel d'émergence humaine
Ils ont fait cela et bien plus encore, comme vous le lirez ci-dessous.
Comme Trump l'a dit, à maintes reprises, les Chinois étaient impliqués.
Le
laboratoire clé des agents pathogènes spéciaux et de la biosécurité,
Institut de virologie de Wuhan, Académie chinoise des sciences, Wuhan,
Chine a fourni le virus de la chauve-souris de Wuhan qui a été utilisé
dans l'étude américaine. Leur nom a été inclus pour cette raison seulement.
COVID
19 était un projet d'armes biologiques de l'armée américaine visant à
fabriquer une maladie causant une pneumonie qu'il serait presque
impossible de vacciner chez les patients de plus de 40 ans.
La preuve est là, faites simplement défiler vers le bas. L'étude a été menée par l'Université de Caroline du Nord et financée par l'USAID / CIA. Ils ont choisi un virus de la chauve-souris chinoise et ont également choisi d'inclure un établissement médical à Wuhan.
Maintenant,
nous savons pourquoi, un écran de fumée du blâme pour un programme avec
la Chine n'avait que peu ou rien à voir, quelque chose de sataniquement
mauvais et purement américain.
En
novembre 2015, une étude a été publiée décrivant la capacité de
produire le virus auquel nous sommes confrontés actuellement. Parmi les nombreuses personnes impliquées, il y avait un laboratoire à Wuhan, en Chine. Il a été répertorié dès le début comme l'un des dizaines de labos, principalement américains, travaillant sur ce projet.
Cependant, un participant clé a été laissé de côté, l'USAID. On
soupçonne, profondément, que l'USAID est une façade pour la recherche
américaine sur la guerre biologique telle que celle menée à Tbilissi, en
Géorgie, et ailleurs, très documentée. Il faut donc ajouter l'USAID au groupe de financement de la recherche de l'arme biologique COVID 19.
Historique des changements
20 novembre 2015
Dans la version de cet article initialement publiée en ligne, les auteurs ont omis de mentionner une source de financement, le financement USAID-EPT-PREDICT d'EcoHealth Alliance, à Z.-LS L'erreur a été corrigée pour les versions imprimée, PDF et HTML de cet article .
Nous
allons maintenant présenter l'article biaisé de Pravda et, ci-dessous,
l'étude réelle prouvant la capacité de produire le COVID 19, prouvant
que ce n'est pas un virus naturel une fois pour toutes.
Quant
à savoir qui a fait quoi, ce n'est pas notre travail mais nous
prouvons, catégoriquement, que lorsqu'un laboratoire chinois est
mentionné, c'est un acteur mineur dans un effort américain, comme décrit
de manière exhaustive ci-dessous.
Cela rend le laboratoire de Wuhan possiblement complice de la guerre biologique.
De
même, lorsque Forbes Magazine et d'autres ont déclaré qu'ils pouvaient
prouver que COVID 19 avait été fabriqué naturellement et qu'ils avaient
bien sûr le même accès que nous, nous soupçonnons qu'ils font partie
d'un effort de désinformation lié à l'USAID et à la bio-guerre.
Le soupçon n'est pas une preuve. La
preuve est la preuve et il y a suffisamment de preuves pour se noyer.
Nos remerciements aux professionnels de la santé américains qui se sont informés auprès de l'armée américaine et de la CIA et qui ont aidé à nous
amener là où nous en sommes, une nation brisée en morceaux.
Pravda.Ru:
Un tel matériel est apparu en 2015 sur le site de la revue scientifique
Natura en 2015. Ensuite, les auteurs ont affirmé qu'après l'avènement
du virus du SRAS (2002-2003) et du syndrome respiratoire du Moyen-Orient
(MERS), les scientifiques étaient au courant du risque de transmission
interspécifique qui conduirait à une épidémie parmi les personnes.
Expérience de laboratoire réussie
Entre autres choses, l'équipe de recherche a étudié les chauves-souris, qui sont les plus grands incubateurs de coronavirus. Néanmoins,
les chauves-souris ne pouvaient pas transmettre le coronavirus aux
humains car elles ne pouvaient pas interagir avec les cellules humaines
avec les récepteurs ACE2.
Le
matériel a également déclaré que les chauves-souris fer à cheval sont
porteuses d'une souche de coronavirus du SRAS qui peut être transmise
aux humains. Il a été nommé virus SHC014-CoV.
Pour mieux étudier ce virus, les scientifiques ont copié le coronavirus et l'ont infecté à des souris de laboratoire. Les
résultats ont montré que le virus est vraiment capable de se lier aux
cellules humaines avec des récepteurs ACE2 et de se multiplier dans les
cellules du système respiratoire.
Dans
les travaux de recherche, il est à noter que le matériel de
laboratoire, les échantillons et l'équipement qui ont été utilisés dans
la recherche ont été obtenus auprès de l'Institut de recherche médicale
de l'armée sur les maladies infectieuses. Bien
qu'il ne soit pas encore possible de dire avec certitude que le virus
qui a été testé sur des souris de laboratoire est le même que le
coronavirus SARS-Cove-2.
Politique de l'OTAN
Cependant, des choses intéressantes peuvent être trouvées dans les documents précédents.
Par exemple:
- Le rapport d'activité 2019 de l'Alliance indique qu'en 2019, la première place de l'Alliance dans la recherche et le développement était occupée par le thème de la protection radiochimique et biologique (29%), déplaçant le problème apparemment le plus pressant de l'Europe - le contre-terrorisme (il s'est avéré être 4 - priorité m).
- Un an plus tôt, en 2018, la situation était exactement le contraire: le terrorisme, comme il se doit, était en premier lieu (28%), et la protection radiochimique et biologique en quatrième (13%).
Comme l'écrit le mouchard de Bruxelles dans la chaîne de télégramme,
«étant donné l'absence de raisons visibles d'un changement aussi
brutal des intérêts scientifiques, il existe deux options et les deux
sont désagréables:
- ou l'OTAN remue maintenant le cinquième point, falsifiant les données pour montrer «et nous nous sommes toujours préparés aux virus, nous sommes modernes»,
- ou même en 2019 dans l'alliance, Dieu me pardonne, ils savaient d'où viendrait le problème.
Oui, la première option est beaucoup plus réelle, mais, voyez-vous, les faits sont surprenants.
Source: Pravda

Recherche originale de 2015 non éditée et terminée
Nature Medicine volume 21 , pages 1508-1513 ( 2015 )
Résumé
L'émergence
du coronavirus du syndrome respiratoire aigu sévère (SRAS-CoV) et du
syndrome respiratoire du Moyen-Orient (MERS) -CoV souligne la menace
d'événements de transmission inter-espèces conduisant à des épidémies
chez l'homme. Nous
examinons ici le potentiel de maladie d'un virus de type SRAS,
SHC014-CoV, qui circule actuellement dans les populations chinoises de
chauves-souris fer à cheval 1 . En utilisant le système de génétique inverse du SRAS-CoV 2
, nous avons généré et caractérisé un virus chimérique exprimant le pic
du coronavirus de chauve-souris SHC014 dans un squelette du SRAS-CoV
adapté à la souris.
Les résultats indiquent que les virus du groupe 2b codant pour le pic SHC014 dans un squelette de type sauvage peuvent utiliser efficacement plusieurs orthologues de l'enzyme II (ACE2) de conversion de l'angiotensine humaine du récepteur du SRAS, se répliquer efficacement dans les cellules des voies respiratoires humaines primaires et atteindre des titres in vitro équivalents à souches épidémiques de SRAS-CoV. De plus, des expériences in vivo démontrent la réplication du virus chimérique dans le poumon de souris avec une pathogenèse notable.
L'évaluation des modalités immunothérapeutiques et prophylactiques basées sur le SRAS a révélé une faible efficacité; les anticorps monoclonaux et l'approche vaccinale n'ont pas réussi à neutraliser et à protéger de l'infection par les CoV en utilisant la nouvelle protéine de pointe.
Sur la base de ces résultats, nous avons dérivé synthétiquement un virus recombinant SHC014 de pleine longueur infectieux et démontrons une réplication virale robuste à la fois in vitro et in vivo . Nos travaux suggèrent un risque potentiel de réémergence du SRAS-CoV à partir de virus circulant actuellement dans les populations de chauves-souris.
Les résultats indiquent que les virus du groupe 2b codant pour le pic SHC014 dans un squelette de type sauvage peuvent utiliser efficacement plusieurs orthologues de l'enzyme II (ACE2) de conversion de l'angiotensine humaine du récepteur du SRAS, se répliquer efficacement dans les cellules des voies respiratoires humaines primaires et atteindre des titres in vitro équivalents à souches épidémiques de SRAS-CoV. De plus, des expériences in vivo démontrent la réplication du virus chimérique dans le poumon de souris avec une pathogenèse notable.
L'évaluation des modalités immunothérapeutiques et prophylactiques basées sur le SRAS a révélé une faible efficacité; les anticorps monoclonaux et l'approche vaccinale n'ont pas réussi à neutraliser et à protéger de l'infection par les CoV en utilisant la nouvelle protéine de pointe.
Sur la base de ces résultats, nous avons dérivé synthétiquement un virus recombinant SHC014 de pleine longueur infectieux et démontrons une réplication virale robuste à la fois in vitro et in vivo . Nos travaux suggèrent un risque potentiel de réémergence du SRAS-CoV à partir de virus circulant actuellement dans les populations de chauves-souris.
L'ESSENTIEL
L'émergence
du SRAS-CoV a inauguré une nouvelle ère dans la transmission
inter-espèces de maladies respiratoires graves, la mondialisation
entraînant une propagation rapide dans le monde et un impact économique
massif 3 , 4
. Depuis lors, plusieurs souches - y compris les souches de grippe A
H5N1, H1N1 et H7N9, et MERS-CoV - ont émergé de populations animales,
causant des maladies, une mortalité et des difficultés économiques
considérables pour les régions touchées 5 . Bien que les mesures de santé publique aient pu arrêter l'épidémie de SRAS-CoV 4
, des études de métagénomique récentes ont identifié des séquences de
virus de type SRAS étroitement apparentés circulant dans les populations
de chauves-souris chinoises qui pourraient constituer une menace future
1 , 6 .
Cependant,
les données de séquence à elles seules fournissent des informations
minimales pour identifier et préparer les futurs virus prépandémiques.
Par conséquent, pour examiner le potentiel d'émergence (c'est-à-dire le
potentiel d'infecter les humains) des CoV de chauve-souris en
circulation, nous
avons construit un virus chimérique codant pour une nouvelle protéine
de pointe CoV zoonotique - à partir de la séquence RsSHC014-CoV qui a
été isolée à partir de fers à cheval chinois 1
—Dans le contexte du squelette adapté à la souris SARS-CoV. Le virus
hybride nous a permis d'évaluer la capacité de la nouvelle protéine de
pointe à provoquer une maladie indépendamment d'autres mutations
adaptatives nécessaires dans son squelette naturel.
En
utilisant cette approche, nous avons caractérisé l'infection par le CoV
médiée par la protéine de pointe SHC014 dans les cellules des voies
respiratoires humaines primaires et in vivo et avons testé l'efficacité des thérapies immunitaires disponibles contre SHC014-CoV. Ensemble, la stratégie traduit les données de métagénomique pour aider à prédire et à se préparer aux futurs virus émergents.
Les
séquences de SHC014 et du RsWIV1-CoV apparenté montrent que ces CoV
sont les plus proches parents des souches épidémiques de SRAS-CoV ( Fig. 1a, b ); cependant,
il existe des différences importantes dans les 14 résidus qui se lient à
l'ACE2 humaine, le récepteur du SRAS-CoV, y compris les cinq qui sont
critiques pour la gamme d'hôtes: Y442, L472, N479, T487 et Y491 (réf. 7 ).
Dans
WIV1, trois de ces résidus diffèrent de la souche épidémique SRAS-CoV
Urbani, mais on ne s'attend pas à ce qu'ils altèrent la liaison à ACE2 (
Fig. 1a, b et tableau supplémentaire 1 supplémentaires
). Ce fait est confirmé par les deux expériences de pseudotypage qui
mesuraient la capacité des lentivirus codant pour les protéines de
pointe WIV1 à pénétrer dans les cellules exprimant l'ACE2 humaine ( figure 1 supplémentaire ) et par des tests de réplication in vitro de WIV1-CoV (réf. 1
). En revanche, 7 des 14 résidus d'interaction ACE2 dans SHC014 sont
différents de ceux dans SARS-CoV, y compris les cinq résidus critiques
pour la gamme d'hôtes ( Fig. 1c supplémentaire et tableau supplémentaire 1 ).
Ces changements, couplés à l'échec des lentivirus pseudotypés exprimant le pic SHC014 à pénétrer dans les cellules ( Fig. 1d supplémentaire
), ont suggéré que le pic SHC014 est incapable de se lier à ACE2
humain. Cependant, des changements similaires dans les souches de
SRAS-CoV apparentées ont été signalés pour permettre la liaison à ACE2 7 , 8 , ce qui suggère que des tests fonctionnels supplémentaires étaient nécessaires pour la vérification.
Par
conséquent, nous avons synthétisé le pic SHC014 dans le contexte du
squelette SRAS-CoV adapté à la souris et compétent pour la réplication
(nous nous référons ci-après au CoV chimérique sous le nom de
SHC014-MA15) afin de maximiser l'opportunité d'études de pathogenèse et
de vaccin chez la souris ( Fig. . 2a
). Malgré les prédictions de la modélisation basée sur la structure et
des expériences de pseudotypage, SHC014-MA15 était viable et répliqué à
des titres élevés dans les cellules Vero ( Fig. 2b supplémentaire
). Semblable au SRAS, SHC014-MA15 nécessitait également une molécule
ACE2 fonctionnelle pour l'entrée et pouvait utiliser des orthologues
ACE2 humains, civettes et chauves-souris ( Fig. 2c, d ) supplémentaires .
Pour tester la capacité de la pointe de SHC014 à l' infection
médiate des voies respiratoires humaines, nous avons examiné la
sensibilité de la lignée cellulaire des voies respiratoires épithéliales
humaines Calu-3 2B4 (réf. 9 ) à l' infection et on a trouvé la réplication SHC014-MA15 robuste, comparable à celle de SARS-CoV Urbani ( Fig. 1c ). Pour
étendre ces résultats, des cultures d'épithélium des voies
respiratoires humaines primaires (AOH) ont été infectées et ont montré
une réplication robuste des deux virus ( Fig. 1d ). Ensemble,
les données confirment la capacité des virus avec le pic SHC014 à
infecter les cellules des voies respiratoires humaines et soulignent la
menace potentielle de transmission inter-espèces de SHC014-CoV.
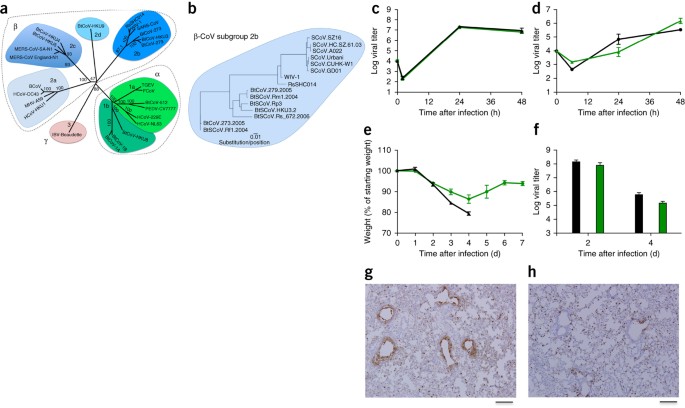
( a
) Les séquences génomiques de pleine longueur des CoV représentatifs
ont été alignées et cartographiées phylogénétiquement comme décrit dans
les méthodes en ligne .
La barre d'échelle représente les substitutions de nucléotides, seul le
support bootstrap supérieur à 70% étant marqué. L'arbre montre les CoV
divisés en trois groupes phylogénétiques distincts, définis comme les
α-CoV, les β-CoV et les γ-CoV. Les clusters de sous-groupes classiques
sont marqués comme 2a, 2b, 2c et 2d pour les β-CoV et comme 1a et 1b
pour les α-CoV. ( b
) Les séquences d'acides aminés des domaines SI des pointes de ß-CoV
représentatifs du groupe 2b, y compris SARS-CoV, ont été alignées et
cartographiées phylogénétiquement. La barre d'échelle représente les
substitutions d'acides aminés. ( c , d) Réplication virale du SRAS-CoV Urbani (noir) et SHC014-MA15 (vert) après infection de cellules Calu-3 2B4 ( c ) ou cultures de cellules AOH à interface air-liquide primaire bien différenciées ( d
) à une multiplicité d'infection (MOI) de 0,01 pour les deux types de
cellules. Des échantillons ont été collectés à des moments individuels
avec des répliques biologiques ( n = 3) pour les expériences Calu-3 et HAE. ( e , f ) Perte de poids ( n = 9 pour SARS-CoV MA15; n = 16 pour SHC014-MA15) ( e ) et réplication virale dans les poumons ( n = 3 pour SARS-CoV MA15; n = 4 pour SHC014- MA15) ( f) Des souris âgées de 10 semaines BALB / c infectées avec 1 x 10 4 pfu du SRAS-CoV MA15 (noir) ou SHC014-MA15 (vert-adapté souris) par voie intranasale (in). ( g, h
) Des images représentatives de coupes pulmonaires colorées pour
l'antigène SRAS-CoV N provenant de souris infectées par SARS-CoV MA15 ( n = 3 souris) ( g ) ou SHC014-MA15 ( n = 4 souris) ( h ) sont présentées. Pour
chaque graphique, la valeur centrale représente la moyenne du groupe et
les barres d'erreur définissent les barres d'échelle sem, 1 mm.
Pour évaluer le rôle du pic SHC014 dans la médiation de l'infection in vivo , nous avons infecté des souris BALB / c âgées de 10 semaines avec 10 4 unités formant plaque (pfu) de SARS-MA15 ou SHC014-MA15 ( Fig. 1e – h
). Les animaux infectés par le SRAS-MA15 ont connu une perte de poids
rapide et une létalité 4 jours après l'infection (dpi); en revanche,
l'infection par SHC014-MA15 a produit une perte de poids substantielle
(10%) mais pas de létalité chez les souris ( figure 1e
). L'examen de la réplication virale a révélé des titres viraux presque
équivalents provenant des poumons de souris infectées par le SRAS-MA15
ou SHC014-MA15 ( figure 1f
). Alors que les poumons des souris infectées par le SRAS-MA15 ont
montré une coloration robuste à la fois dans les bronchioles terminales
et le parenchyme pulmonaire 2 dpi ( Fig. 1g), ceux des souris infectées par SHC014-MA15 ont montré une coloration antigénique des voies respiratoires réduite ( Fig. 1h
); en revanche, aucun déficit de coloration de l'antigène n'a été
observé dans le parenchyme ou dans le score histologique global,
suggérant une infection différentielle du tissu pulmonaire pour
SHC014-MA15 ( tableau supplémentaire 2
). Nous avons ensuite analysé l'infection chez des animaux âgés (âgés
de 12 mois) plus sensibles. Les animaux infectés par le SRAS-MA15 ont
rapidement perdu du poids et ont succombé à l'infection ( Fig. 3a, b ) supplémentaires
. SHC014-MA15 a induit une perte de poids robuste et soutenue, mais a
une létalité minimale. Les tendances de l'histologie et des schémas de
coloration de l'antigène que nous avons observés chez les jeunes souris
ont été conservées chez les animaux plus âgés ( tableau supplémentaire 3).
Nous avons exclu la possibilité que SHC014-MA15 médie l'infection par
un récepteur alternatif sur la base d'expériences utilisant des souris Ace2 - / - , qui n'ont pas montré de perte de poids ou de coloration de l'antigène après l'infection par SHC014-MA15 ( Fig. 4a, b et supplémentaire) Tableau 2
). Ensemble, les données indiquent que les virus avec le pic SHC014
sont capables d'induire une perte de poids chez la souris dans le
contexte d'un squelette virulent du CoV.
Compte tenu de l'efficacité préclinique des thérapies par anticorps monoclonaux Ebola, telles que ZMApp 10
, nous avons ensuite cherché à déterminer l'efficacité des anticorps
monoclonaux contre le SRAS-CoV contre l'infection par SHC014-MA15.
Quatre anticorps monoclonaux humains largement neutralisants ciblant la
protéine de pointe du SRAS-CoV avaient déjà été rapportés et sont des
réactifs probables pour l'immunothérapie 11 , 12 , 13
. Nous avons examiné l'effet de ces anticorps sur la réplication virale
(exprimé en pourcentage d'inhibition de la réplication virale) et
constaté que, alors que le SARS-CoV Urbani de type sauvage était
fortement neutralisé par les quatre anticorps à des concentrations
d'anticorps relativement faibles ( Fig. 2a – d), la neutralisation a varié pour SHC014-MA15. Fm6, un anticorps généré par présentation de phage et mutants d'échappement 11 , 12 , atteint uniquement des niveaux de fond d'inhibition de la réplication de SHC014-MA15 ( Fig. 2a ). De même, les anticorps 230.15 et 227.14, qui étaient dérivés de cellules B mémoire de patients infectés par le SRAS-CoV 13 , ont également échoué à bloquer la réplication de SHC014-MA15 ( Fig. 2b, c). Pour
les trois anticorps, les différences entre les séquences d'acides
aminés du pic du SRAS et du SHC014 correspondaient à des changements de
résidus directs ou adjacents trouvés dans les mutants d'échappement du
SARS-CoV (fm6 N479R; 230,15 L443V; 227,14 K390Q / E), ce qui explique
probablement l'absence des anticorps 'activité neutralisante contre
SHC014. Enfin, l'anticorps
monoclonal 109.8 a pu atteindre 50% de neutralisation de SHC014-MA15,
mais uniquement à des concentrations élevées (10 µg / ml) ( Fig. 2d ). Ensemble,
les résultats démontrent que les anticorps neutralisant largement
contre le SARS-CoV peuvent n'avoir qu'une efficacité marginale contre
les souches émergentes de CoV de type SRAS telles que SHC014.
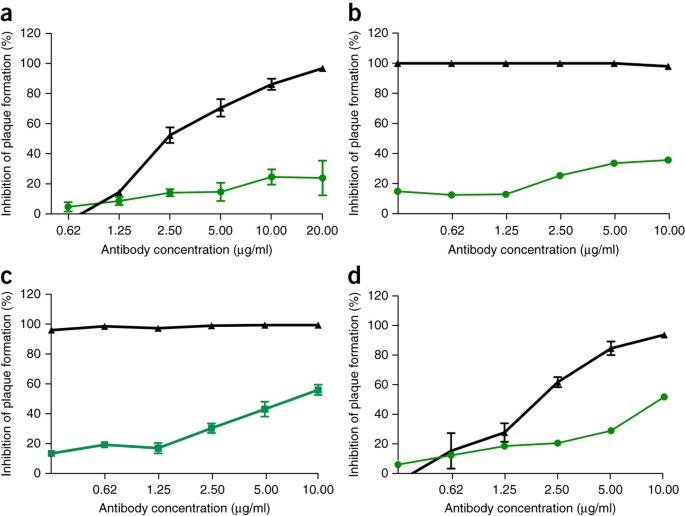
( a - d
) Essais de neutralisation évaluant l'efficacité (mesurée comme une
réduction du nombre de plaques) d'un panel d'anticorps monoclonaux, qui
ont tous été initialement générés contre l'épidémie de SRAS-CoV, contre
l'infection des cellules Vero par SRAS-CoV Urbani (noir ) ou SHC014-MA15
(vert). Les anticorps testés étaient fm6 ( n = 3 pour Urbani; n = 5 pour SHC014-MA15) 11 , 12 ( a ), 230,15 ( n = 3 pour Urbani; n = 2 pour SHC014-MA15) ( b ), 227,15 ( n = 3 pour Urbani; n = 5 pour SHC014-MA15) ( c ) et 109,8 ( n = 3 pour Urbani; n = 2 pour SHC014-MA15) 13 ( d ). Chaque
point de données représente la moyenne du groupe et les barres d'erreur
définissent le sem. Notez que les barres d'erreur dans les cellules
Vero infectées par SRAS-CoV Urbani dans b , c sont recouvertes par les symboles et ne sont pas visibles.
Pour
évaluer l'efficacité des vaccins existants contre l'infection par
SHC014-MA15, nous avons vacciné des souris âgées avec le SRAS-CoV (DIV)
entier double inactivé. Des travaux antérieurs ont montré que DIV
pouvait neutraliser et protéger les jeunes souris contre une provocation
avec un virus homologue 14
; cependant, le vaccin n'a pas réussi à protéger les animaux âgés chez
lesquels une pathologie immunitaire augmentée a également été observée,
ce qui indique la possibilité que les animaux soient lésés en raison de
la vaccination 15
. Ici, nous avons constaté que DIV ne fournissait pas de protection
contre la provocation avec SHC014-MA15 en ce qui concerne la perte de
poids ou le titre viral ( Fig. 5a, b ) supplémentaire . Conformément à un rapport précédent avec d'autres groupes hétérologues 2b CoVs 15, le sérum de souris âgées vaccinées par DIV n'a pas réussi non plus à neutraliser SHC014-MA15 ( figure 5c supplémentaire ). Notamment, la vaccination DIV a entraîné une pathologie immunitaire robuste ( tableau supplémentaire 4 ) et une éosinophilie ( figure supplémentaire 5d-f
). Ensemble, ces résultats confirment que le vaccin DIV ne serait pas
protecteur contre l'infection par SHC014 et pourrait éventuellement
augmenter la maladie dans le groupe vacciné âgé.
Contrairement
à la vaccination de souris avec DIV, l'utilisation de SHC014-MA15 comme
vaccin vivant atténué a montré une protection croisée potentielle
contre la provocation avec le SRAS-CoV, mais les résultats comportent
des mises en garde importantes. Nous avons infecté les jeunes souris
avec 10 4
pfu de SHC014-MA15 et les avons observées pendant 28 jours. Nous avons
ensuite provoqué les souris avec le SRAS-MA15 au jour 29 ( Fig. 6a supplémentaire
). L'infection antérieure des souris avec la dose élevée de SHC014-MA15
conférait une protection contre la provocation avec une dose létale de
SRAS-MA15, bien qu'il n'y ait eu qu'une réponse de neutralisation du
SRAS-CoV minimale des antisérums provoqués 28 jours après l'infection
par SHC014-MA15 ( Supplémentaire Fig.6b,
1: 200). En l'absence d'un rappel d'antigène secondaire, 28 dpi
représente le pic attendu des titres d'anticorps et implique qu'il y
aura une protection diminuée contre le SRAS-CoV au fil du temps 16 , 17
. Des résultats similaires montrant une protection contre une
provocation avec une dose létale de SRAS-CoV ont été observés chez des
souris BALB / c âgées en ce qui concerne la perte de poids et la
réplication virale ( Fig. 6c, d ) supplémentaires . Cependant, la dose d'infection SHC014-MA15 de 10 4 pfu a induit une perte de poids et une létalité> 10% chez certains animaux âgés ( Fig.1 et Fig.3).
Nous avons constaté que la vaccination avec une dose plus faible de
SHC014-MA15 (100 pfu), n'a pas induit de perte de poids, mais elle a
également échoué à protéger les animaux âgés contre une dose létale du
SRAS-MA15 ( figure supplémentaire 6e, f
). Ensemble, les données suggèrent que la provocation SHC014-MA15 peut
conférer une protection croisée contre le SRAS-CoV par le biais
d'épitopes conservés, mais la dose requise induit une pathogenèse et
empêche l'utilisation comme vaccin atténué.
Après
avoir établi que le pic SHC014 a la capacité de médier l'infection des
cellules humaines et de provoquer une maladie chez la souris, nous avons
ensuite synthétisé un clone infectieux SHC014-CoV complet basé sur
l'approche utilisée pour le SRAS-CoV ( Fig.3a ) 2 . La réplication dans les cellules Vero n'a révélé aucun déficit pour SHC014-CoV par rapport à celui pour SRAS-CoV ( figure 3b ); cependant, SHC014-CoV était significativement atténué ( P <0 24="" 48="" apr="" cultures="" d="" dans="" et="" fois="" font="" h="" infection="" l="" la="" les="" primaires="" s="">Fig. 3c ). L' infection in vivo
de souris n'a démontré aucune perte de poids significative, mais a
montré une réplication virale réduite dans les poumons de l'infection
SHC014-CoV pleine longueur, par rapport au SRAS-CoV Urbani ( Fig. 3d, e). Ensemble,
les résultats établissent la viabilité du SHC014-CoV pleine longueur,
mais suggèrent qu'une adaptation supplémentaire est nécessaire pour que
sa réplication soit équivalente à celle du SRAS-CoV épidémique dans les
cellules respiratoires humaines et chez la souris.
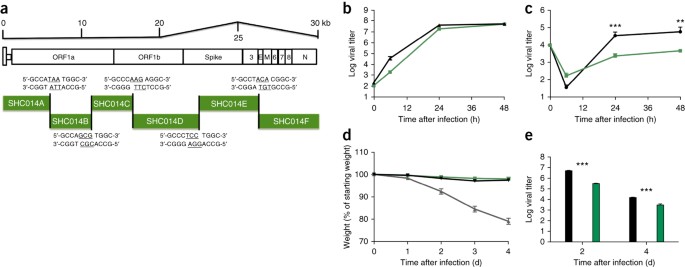
( a
) Schéma du clone moléculaire SHC014-CoV, qui a été synthétisé sous
forme de six ADNc contigus (désignés SHC014A, SHC014B, SHC014C, SHC014D,
SHC014E et SHC014F) flanqués de sites BglI uniques qui ont permis
l'assemblage dirigé de l'ADNc de pleine longueur exprimant cadres de
lecture ouverts (pour 1a, 1b, spike, 3, enveloppe, matrice, 6–8 et
nucléocapside). Les nucléotides soulignés représentent les séquences en
surplomb formées après le clivage par l'enzyme de restriction. ( b , c ) Réplication virale du SRAS-CoV Urbani (noir) ou SHC014-CoV (vert) après infection de cellules Vero ( b ) ou cultures de cellules AOH à interface air-liquide primaire bien différenciées ( c) à un MOI de 0,01. Des échantillons ont été collectés à des moments individuels avec des répliques biologiques ( n = 3) pour chaque groupe. Les données représentent une expérience pour les cellules Vero et HAE. ( d , e ) Perte de poids ( n = 3 pour SRAS-CoV MA15, n = 7 pour SHC014-CoV; n = 6 pour SRAS-Urbani) ( d ) et réplication virale dans les poumons ( n = 3 pour SRAS-Urbani et SHC014-CoV) ( e ) de souris BALB / c âgées de 10 semaines infectées avec 1 × 10 5
pfu de SARS-CoV MA15 (gris), SHC014-CoV (vert) ou SARS-CoV Urbani
(noir) via le en route. Chaque point de données représente la moyenne du
groupe et les barres d'erreur définissent le sem **P <0 et="" font="">P <0 en="" font="" le="" utilisant="">test t de Student bilatéral des points de temps individuels.
Lors
de l'épidémie de SRAS-CoV, des liens ont été rapidement établis entre
les civettes de palmiers et les souches de CoV détectées chez l'homme 4 . Fort de ce constat, le paradigme de l' émergence commune soutient que l' épidémie de SRAS-CoV origine un virus de la chauve - souris, a sauté à civettes, et les changements incorporés dans le domaine de liaison au récepteur (RBD) pour améliorer la liaison à civette Ace2 (réf. 18
). Une exposition ultérieure à des personnes sur les marchés d'animaux
vivants a permis l'infection humaine par la souche de civette, qui, à
son tour, s'est adaptée pour devenir la souche épidémique ( Fig. 4a
). Cependant, l'analyse phylogénétique suggère que les premières
souches du SRAS humain semblent plus étroitement liées aux souches de
chauves-souris qu'aux souches de civettes 18. Par
conséquent, un deuxième paradigme soutient que la transmission directe
entre les chauves-souris et l'homme a déclenché l'émergence du SRAS-CoV
et que les civettes de palmiers ont servi d'hôte secondaire et de
réservoir pour une infection continue ( Fig. 4b ) 19 . Pour
les deux paradigmes, l'adaptation des pics chez un hôte secondaire est
considérée comme une nécessité, la plupart des mutations devant se
produire dans le RBD, facilitant ainsi une infection améliorée. Les
deux théories impliquent que les pools de CoV de chauves-souris sont
limités et que les mutations de la gamme d'hôtes sont à la fois
aléatoires et rares, ce qui réduit la probabilité d'événements
d'émergence futurs chez l'homme.
Les souches de coronavirus sont maintenues dans des pools de quasi-espèces circulant dans les populations de chauves-souris. ( a , b
) Les théories traditionnelles d'émergence du SRAS-CoV postulent que
les mutants de la gamme d'hôtes (cercle rouge) représentent des
occurrences aléatoires et rares qui permettent l'infection d'hôtes
alternatifs. Le paradigme de l'hôte secondaire ( a
) fait valoir qu'un hôte non humain est infecté par un virus
progéniteur de chauve-souris et, par adaptation, facilite la
transmission aux humains; la réplication ultérieure chez l'homme conduit
à la souche virale épidémique. Le paradigme direct ( b) suggère que la transmission se produit entre les chauves-souris et les humains sans avoir besoin d'un hôte intermédiaire; la
sélection se produit alors dans la population humaine avec des virus
étroitement apparentés se répliquant dans un hôte secondaire, permettant
une persistance virale continue et une adaptation dans les deux. ( c
) Les données des virus chimériques de type SRAS font valoir que les
pools de quasi-espèces maintiennent plusieurs virus capables d'infecter
les cellules humaines sans nécessiter de mutations (cercles rouges). Bien
que des adaptations chez des hôtes secondaires ou humains puissent être
nécessaires pour l'émergence d'une épidémie, si des virus contenant des
pics SHC014 sont recombinés avec des squelettes virulents de CoV
(cercles avec des contours verts), une maladie épidémique peut en
résulter chez l'homme. Les données existantes prennent en charge les éléments des trois paradigmes.
Bien
que notre étude n'invalide pas les autres voies d'émergence, elle
plaide pour un troisième paradigme dans lequel les pools de CoV de
chauves-souris en circulation maintiennent des protéines de pointe `` en
équilibre '' capables d'infecter les humains sans mutation ni
adaptation ( Fig.4c
). Cette hypothèse est illustrée par la capacité d'un virus chimérique
contenant le pic SHC014 dans un squelette SRAS-CoV à provoquer une
infection robuste dans les cultures des voies respiratoires humaines et
chez les souris sans adaptation RBD.
Couplés à l'observation de squelettes CoV pathogènes précédemment identifiés 3 , 20
, nos résultats suggèrent que les matériaux de départ nécessaires pour
les souches émergentes de type SRAS circulent actuellement dans les
réservoirs animaux. Notamment, bien que SHC014-CoV pleine longueur
nécessite probablement une adaptation supplémentaire du squelette pour
médier la maladie humaine, les événements de recombinaison à haute
fréquence documentés dans les familles de CoV soulignent la possibilité
d'une émergence future et la nécessité d'une préparation supplémentaire.
À
ce jour, les criblages génomiques des populations animales ont
principalement été utilisés pour identifier de nouveaux virus dans les
contextes d'épidémie 21
. L'approche ici étend ces ensembles de données pour examiner les
questions d'émergence virale et d'efficacité thérapeutique. Nous
considérons les virus avec le pic SHC014 comme une menace potentielle en
raison de leur capacité à se répliquer dans les cultures primaires des
voies respiratoires humaines, le meilleur modèle disponible pour la
maladie humaine. De plus, la pathogenèse observée chez la souris indique
une capacité des virus contenant SHC014 à provoquer une maladie chez
des modèles mammifères, sans adaptation RBD.
Notamment,
le tropisme différentiel dans le poumon par rapport à celui du
SRAS-MA15 et l'atténuation du SHC014-CoV de pleine longueur dans les
cultures d'AOH par rapport au SRAS-CoV Urbani suggèrent que des facteurs
au-delà de la liaison ACE2 - y compris la processivité des pics, la
biodisponibilité des récepteurs ou l'antagonisme des réponses
immunitaires de l'hôte - peuvent contribuer à l'émergence. Cependant,
des tests supplémentaires sur des primates non humains sont nécessaires
pour traduire ces résultats en potentiel pathogène chez l'homme.
Surtout,
l'échec des thérapies disponibles définit un besoin critique pour une
étude plus approfondie et pour le développement de traitements. Avec
ces connaissances, des programmes de surveillance, des réactifs de
diagnostic et des traitements efficaces peuvent être produits qui
protègent contre l'émergence de CoV spécifiques du groupe 2b, tels que
SHC014, et ceux-ci peuvent être appliqués à d'autres branches de CoV qui
conservent des pools également hétérogènes.
En
plus d'offrir une préparation contre les futurs virus émergents, cette
approche doit être envisagée dans le contexte de la pause imposée par le
gouvernement américain sur les études sur le gain de fonction (GOF) 22 .
Sur la base des modèles d'émergence précédents ( Fig. 4a, b ), la création de virus chimériques tels que SHC014-MA15 ne devrait pas augmenter la pathogénicité. Bien
que SHC014-MA15 soit atténué par rapport à son SRAS-CoV parental adapté
à la souris, des études similaires examinant la pathogénicité des CoV
avec le pic Urbani de type sauvage dans le squelette MA15 n'ont montré
aucune perte de poids chez la souris et une réplication virale réduite 23 . Ainsi, par rapport au pic Urbani – MA15 CoV, SHC014-MA15 montre un gain en pathogenèse ( Fig. 1 ).
Sur
la base de ces résultats, les comités d'examen scientifique peuvent
juger des études similaires construisant des virus chimériques basés sur
des souches en circulation trop risquées à poursuivre, car une
pathogénicité accrue dans les modèles mammifères ne peut être exclue.
Couplée
aux restrictions sur les souches adaptées à la souris et au
développement d'anticorps monoclonaux utilisant des mutants
d'échappement, la recherche sur l'émergence du CoV et l'efficacité
thérapeutique pourrait être sévèrement limitée à l'avenir. Ensemble, ces
données et restrictions représentent un carrefour des préoccupations de
recherche du GOF; le potentiel de préparation et d'atténuation des
épidémies futures doit être mis en balance avec le risque de créer des
agents pathogènes plus dangereux. Lors de l'élaboration de politiques à
l'avenir, il est important de considérer la valeur des données générées
par ces études et de déterminer si ces types d'études sur les virus
chimériques justifient une enquête plus approfondie par rapport aux
risques inhérents impliqués.
Dans
l'ensemble, notre approche a utilisé des données de métagénomique pour
identifier une menace potentielle posée par le CoV SHC014 de type SRAS
de chauve-souris en circulation. En
raison de la capacité des virus SHC014 chimériques à se répliquer dans
les cultures des voies respiratoires humaines, à provoquer une
pathogenèse in vivo
et à échapper aux thérapies actuelles, il existe un besoin à la fois
de surveillance et de thérapies améliorées contre les virus de type SRAS
en circulation. Notre
approche débloque également l'utilisation des données de métagénomique
pour prédire l'émergence virale et appliquer ces connaissances pour se
préparer à traiter les futures infections virales émergentes.
Méthodes
Virus, cellules, infections in vitro et tests de plaque
Le
SARS-CoV de type sauvage (Urbani), le SARS-CoV (MA15) adapté à la
souris et les CoV chimériques semblables au SRAS ont été cultivés sur
des cellules Vero E6 (obtenues auprès de l'Institut de recherche
médicale de l'armée des États-Unis sur les maladies infectieuses),
cultivées dans le Eagle modifié de Dulbecco. milieu (DMEM) (Gibco, CA)
et 5% de sérum de clone foetal (FCS) (Hyclone, South Logan, UT) avec un
antibiotique / antimycotique (Gibco, Carlsbad, CA). Des cellules DBT
(laboratoire Baric, source inconnue) exprimant des orthologues ACE2 ont été précédemment décrites pour l'homme et la civette; La séquence bat Ace2 était basée sur celle de Rhinolophus leschenaulti , et les cellules DBT exprimant bat Ace2 ont été établies comme décrit précédemment 8 .
Les
expériences de pseudotypage étaient similaires à celles utilisant un
pseudovirus basé sur le VIH, préparé comme décrit précédemment 10 , et examinés sur des cellules HeLa (Wuhan Institute of Virology) qui exprimaient des orthologues ACE2
. Les cellules HeLa ont été cultivées dans un milieu essentiel minimal
(MEM) (Gibco, CA) supplémenté avec 10% de FCS (Gibco, CA) comme décrit
précédemment 24 .
Les
courbes de croissance dans Vero E6, DBT, Calu-3 2B4 et les cellules
épithéliales des voies aériennes humaines primaires ont été réalisées
comme décrit précédemment 8 , 25
. Aucun des stocks de lignées cellulaires de travail n'a été
authentifié ou testé récemment pour le mycoplasme, bien que les stocks
de semences d'origine utilisés pour créer les stocks de travail soient
exempts de contamination. Les poumons humains pour les cultures d'AOH
ont été achetés dans le cadre de protocoles approuvés par le Conseil
d'examen institutionnel de l'Université de Caroline du Nord à Chapel
Hill. Les cultures d'AOH représentent un épithélium des voies
respiratoires humaines hautement différencié contenant des cellules
épithéliales ciliées et non ciliées ainsi que des cellules caliciformes.
Les cultures sont également cultivées sur une interface air-liquide
pendant plusieurs semaines avant utilisation, comme décrit précédemment 26 .
En
bref, les cellules ont été lavées avec du PBS et inoculées avec le
virus ou diluées de façon simulée dans du PBS pendant 40 minutes à 37 °
C. Après inoculation, les cellules ont été lavées trois fois et un milieu frais a été ajouté pour signifier le temps «0». Au moins trois réplicats biologiques ont été récoltés à chaque instant décrit. Aucun aveugle n'a été utilisé dans les collections d'échantillons et les échantillons n'ont pas été randomisés. Toutes
les cultures de virus ont été réalisées dans un laboratoire de niveau
de sécurité biologique (BSL) 3 avec des ventilateurs redondants dans les
enceintes de sécurité biologique, comme décrit précédemment par notre
groupe 2 . Tout
le personnel portait des respirateurs à épuration d'air motorisés
(Breathe Easy, 3M) avec des combinaisons Tyvek, des tabliers et des
chaussons et portait des gants doubles.
Clustering de séquences et modélisation structurelle.
Les
séquences génomiques complètes et les séquences d'acides aminés des
domaines S1 du pic de CoV représentatifs ont été téléchargées à partir
de Genbank ou Pathosystems Resource Integration Center (PATRIC),
alignées avec ClustalX et comparées phylogénétiquement en utilisant
l'estimation du maximum de vraisemblance à l'aide de 100 bootstraps ou
par en utilisant le package PhyML
, respectivement. L'arbre a été généré en utilisant le maximum de
vraisemblance avec le package PhyML. La barre d'échelle représente les
substitutions de nucléotides. Seuls les nœuds avec un support d'amorçage
supérieur à 70% sont étiquetés.
L'arbre
montre que les CoV sont divisés en trois groupes phylogénétiques
distincts définis comme les α-CoV, les β-CoV et les γ-CoV. Les clusters de sous-groupes classiques sont marqués comme 2a, 2b, 2c et 2d pour les β-CoV et 1a et 1b pour les α-CoV. Des
modèles structurels ont été générés à l'aide de Modeller (Max Planck
Institute Bioinformatics Toolkit) pour générer des modèles d'homologie
pour SHC014 et Rs3367 du SRAS RBD en complexe avec ACE2 basé sur la
structure cristalline 2AJF (Protein Data Bank). Les modèles d'homologie ont été visualisés et manipulés dans MacPyMol (version 1.3).
Construction de virus chimériques de type SRAS.
Les
virus de type sauvage et chimériques sont dérivés soit du SARS-CoV
Urbani, soit du clone infectieux (ic) adapté à la souris correspondant
(SARS-CoV MA15) comme décrit précédemment 27
. Les plasmides contenant des séquences de pic pour SHC014 ont été
extraits par digestion de restriction et ligaturés dans les plasmides E
et F du clone infectieux MA15. Le clone a été conçu et acheté auprès de
Bio Basic sous forme de six ADNc contigus en utilisant des séquences
publiées flanquées de sites d'endonucléase de restriction de classe II
uniques (BglI). Ensuite, des plasmides contenant des fragments de génome
de SARS-CoV chimériques de type sauvage et de SHC014-CoV ont été
amplifiés, excisés, ligaturés et purifiés.
Des réactions de transcription in vitro
ont ensuite été effectuées pour synthétiser de l'ARN génomique de
pleine longueur, qui a été transfecté dans des cellules Vero E6 comme
décrit précédemment 2 . Le milieu des cellules transfectées a été récolté et a servi de stocks de graines pour des expériences ultérieures. Les virus chimériques et de pleine longueur ont été confirmés par analyse de séquence avant utilisation dans ces études. La
construction synthétique du mutant chimérique SHC014-CoV de pleine
longueur a été approuvée par le comité de biosécurité institutionnelle
de l'Université de Caroline du Nord et le comité de recherche à double
usage sur les préoccupations.
Déclaration d'éthique
Cette
étude a été réalisée conformément aux recommandations pour le soin et
l'utilisation des animaux par l'Office of Laboratory Animal Welfare
(OLAW), NIH. Le Comité
institutionnel de soin et d'utilisation des animaux (IACUC) de
l'Université de Caroline du Nord à Chapel Hill (UNC, numéro de permis
A-3410-01) a approuvé le protocole d'étude sur les animaux (IACUC #
13-033) utilisé dans ces études.
Souris et infection in vivo
Des
souris BALB / cAnNHsD femelles, âgées de 10 semaines et 12 mois ont été
commandées aux laboratoires Harlan. Les infections de souris ont été
effectuées comme décrit précédemment 20
. En bref, les animaux ont été amenés dans un laboratoire BSL3 et
autorisés à s'acclimater pendant 1 semaine avant l'infection. Pour
l'infection et la vaccination contre le virus vivant atténué, les souris
ont été anesthésiées avec un mélange de kétamine et de xylazine et
infectées par voie intranasale, lorsqu'elles ont été provoquées, avec 50
pi de solution saline tamponnée au phosphate (PBS) ou de virus dilué
avec trois ou quatre souris par point de temps, par groupe d'infection
par dose comme décrit dans les légendes de la figure.
Pour
les souris individuelles, les notations d'infection, y compris l'échec
de l'inhalation de la dose entière, la formation de bulles d'inoculum
par le nez ou l'infection par la bouche, peuvent avoir conduit à
l'exclusion des données sur les souris à la discrétion du chercheur;
post-infection, aucun autre critère d'exclusion ou d'inclusion préétabli
n'est défini. Aucune mise en aveugle n'a été utilisée dans aucune
expérimentation animale, et les animaux n'ont pas été randomisés. Pour
la vaccination, des souris jeunes et âgées ont été vaccinées par
injection de coussinet plantaire avec un volume de 20 μl de 0,2 μg de
vaccin contre le SRAS-CoV double inactivé avec de l'alun ou du PBS
simulé; les souris ont ensuite reçu un rappel avec le même régime 22
jours plus tard et ont été stimulées 21 jours après. Pour tous les
groupes, selon le protocole, les animaux ont été surveillés
quotidiennement pour détecter les signes cliniques de maladie (courbure,
fourrure ébouriffée et activité réduite) pendant la durée de
l'expérience.La perte de poids a été surveillée quotidiennement pendant
les 7 premiers jours, après quoi la surveillance du poids s'est
poursuivie jusqu'à ce que les animaux récupèrent leur poids de départ
initial ou affichent un gain de poids en continu pendant 3 jours.
Toutes
les souris qui ont perdu plus de 20% de leur poids corporel de départ
ont été nourries au sol et surveillées plusieurs fois par jour tant
qu'elles étaient sous le seuil de 20%. Les souris qui ont perdu plus de 30% de leur poids corporel de départ ont été immédiatement sacrifiées selon le protocole. Toute
souris jugée moribonde ou peu susceptible de récupérer a également été
sacrifiée humainement à la discrétion du chercheur. L'euthanasie a été réalisée en utilisant une surdose d'isoflurane et la mort a été confirmée par luxation cervicale. Toutes
les études sur les souris ont été réalisées à l'Université de Caroline
du Nord (Animal Welfare Assurance # A3410-01) en utilisant des
protocoles approuvés par le Comité institutionnel de soin et
d'utilisation des animaux de l'UNC (IACUC).
Analyse histologique.
Le poumon gauche a été prélevé et immergé dans du formol tamponné à 10% (Fisher) sans gonflage pendant 1 semaine. Les
tissus ont été inclus dans de la paraffine et des coupes de 5 μm ont
été préparées par le centre d'histopathologie UNC Lineberger
Comprehensive Cancer Center. Pour
déterminer l'étendue de la coloration de l'antigène, les coupes ont été
colorées pour l'antigène viral en utilisant un anticorps
anti-nucléocapside polyclonal SARS-CoV disponible dans le commerce
(Imgenex) et notées en aveugle par pour la coloration des voies
respiratoires et du parenchyme comme décrit précédemment 20 . Les images ont été capturées à l'aide d'un microscope Olympus BX41 avec une caméra Olympus DP71.
Tests de neutralisation virale.
Des
dosages de titre de neutralisation de réduction de plaque ont été
réalisés avec des anticorps préalablement caractérisés contre le
SRAS-CoV, comme décrit précédemment 11 ,
. En
bref, des anticorps neutralisants ou du sérum ont été dilués en série
deux fois et incubés avec 100 pfu des différentes souches infectieuses
de clone de SARS-CoV pendant 1 h à 37 ° C. Le virus et d' anticorps ont ensuite été ajoutés à une plaque à 6 puits avec 5 x 10 5 cellules Vero E6 / puits avec de multiples répétitions ( n ≥ 2). Après une incubation d'une heure à 37 ° C, les cellules ont été recouvertes de 3 ml d'agarose à 0,8% dans un milieu. Les
plaques ont été incubées pendant 2 jours à 37 ° C, colorées avec du
rouge neutre pendant 3 h et les plaques ont été comptées. Le
pourcentage de réduction de plaque a été calculé comme suit: (1 -
(nombre de plaques avec un anticorps / nombre de plaques sans
anticorps)) x 100.
analyses statistiques
Toutes
les expériences ont été menées en opposant deux groupes expérimentaux
(soit deux virus, soit des cohortes vaccinées et non vaccinées). Par conséquent, des différences significatives dans le titre viral et la notation histologique ont été déterminées par un test t de Student bilatéral à des moments individuels. Les données étaient normalement distribuées dans chaque groupe comparé et avaient une variance similaire.
Biosécurité
Les
études rapportées ont été lancées après que le comité de biosécurité
institutionnel de l'Université de Caroline du Nord a approuvé le
protocole expérimental (Titre du projet: Génération de clones infectieux
de CoV de chauve-souris de type SRAS; ID du plan de sécurité du
laboratoire: 20145741; ID de l'annexe G: 12279).
Ces études ont été lancées avant la suspension
du financement de la recherche sur le processus délibératif du
gouvernement américain sur une sélection de recherche sur le gain de
fonction impliquant des virus de la grippe, du MERS et du SRAS
. Ce document a été révisé par l'agence de financement, le NIH. La
poursuite de ces études a été demandée, et cela a été approuvé par le
NIH.
SARS-CoV
est un agent de sélection. Tous les travaux relatifs à ces études ont
été effectués avec des procédures d'exploitation normalisées (SOP) et
des conditions de sécurité approuvées pour le SRAS-CoV, les MERs-CoV et
d'autres CoV connexes. Nos installations institutionnelles CoV BSL3 ont
été conçues pour se conformer aux exigences de sécurité recommandées par
le Biosafety in Microbiological and Biomedical Laboratories (BMBL), le
US Department of Health and Human Services, le Public Health Service,
les Centers for Disease Control (CDC ) et le NIH. Des plans de sécurité
du laboratoire ont été soumis et l'installation a été approuvée pour
utilisation par le Département de la santé et de la sécurité
environnementales (EHS) de l'UNC et le CDC. Un accès par carte
électronique est requis pour entrer dans l'établissement.
Tous
les travailleurs ont été formés par EHS pour utiliser en toute sécurité
des respirateurs à épuration d'air motorisés (PAPR), et des habitudes
de travail appropriées dans une installation BSL3 et des plans de
surveillance médicale active sont en place. Nos installations CoV BSL3
contiennent des ventilateurs redondants, une alimentation de secours
pour les ventilateurs et les armoires et congélateurs de sécurité
biologique, et nos installations peuvent accueillir des racks de souris
SealSafe. Les matériaux classés comme agents BSL3 sont constitués de
SARS-CoV, de souches précurseurs de chauve-souris CoV, de MERS-CoV et de
mutants dérivés de ces agents pathogènes. Au sein des installations
BSL3, l'expérimentation d'un virus infectieux est réalisée dans une
armoire de biosécurité (BSC) de classe II certifiée.
Tous
les membres du personnel portent des gommages, des combinaisons et des
tabliers Tyvek, des PAPR et des couvre-chaussures, et leurs mains
portent des gants doubles. Les
utilisateurs de BSL3 sont soumis à un plan de surveillance médicale
surveillé par la Clinique de santé au travail des employés de
l'université (UEOHC), qui comprend une vaccination annuelle contre la
grippe physique et annuelle et la déclaration obligatoire de tout
symptôme associé à une infection au CoV pendant les périodes de travail
dans le BSL3. Tous les
utilisateurs de BSL3 sont formés à la gestion de l'exposition et aux
protocoles de notification, sont préparés à l'auto-quarantaine et ont
été formés pour être livrés en toute sécurité à un service local de
gestion des maladies infectieuses en cas d'urgence. Tous
les événements d'exposition potentiels sont signalés et étudiés par
l'EHS et l'UEOHC, avec des rapports déposés à la fois au CDC et au NIH.
Codes d'adhésion
Les accessions
Banque de données sur les protéines
Historique des changements
20 novembre 2015
Dans
la version de cet article initialement publiée en ligne, les auteurs
ont omis de mentionner une source de financement, le financement
USAID-EPT-PREDICT d'EcoHealth Alliance, à Z.-LS L'erreur a été corrigée
pour les versions imprimée, PDF et HTML de cet article .
Références
- Ge, XY et coll. Isolement et caractérisation d'un coronavirus de type SRAS de chauve-souris qui utilise le récepteur ACE2. Nature 503 , 535–538 (2013).
- Yount, B. et coll. Génétique inverse avec un ADNc infectieux complet du coronavirus du syndrome respiratoire aigu sévère. Proc. Natl. Acad. Sci. USA 100 , 12995–13000 (2003).
- Becker, MM et al. Le coronavirus synthétique de type SRAS de chauve-souris recombinant est infectieux dans les cellules cultivées et chez la souris. Proc. Natl. Acad. Sci. USA 105 , 19944–19949 (2008).
- Peiris, JS, Guan, Y. & Yuen, KY Syndrome respiratoire aigu sévère. Nat. Med. 10 , S88-S97 (2004).
- Al-Tawfiq, JA et al. Surveillance des virus respiratoires émergents. Lancet Infect. Dis. 14 , 992-1000 (2014).
- He, B. et coll. Identification de divers alphacoronavirus et caractérisation génomique d'un nouveau coronavirus de type syndrome respiratoire aigu sévère chez les chauves-souris en Chine. J. Virol. 88 , 7070-7082 (2014).
- Li, F. Reconnaissance des récepteurs et infections interspécifiques du coronavirus du SRAS. Antiviral Res. 100 , 246–254 (2013).
- Sheahan, T. et coll. Mécanismes de l'expansion de la gamme d'hôtes du coronavirus du syndrome respiratoire aigu sévère zoonotique dans l'épithélium des voies respiratoires humaines. J. Virol. 82 , 2274-2285 (2008).
- Yoshikawa, T. et coll. Réponses immunitaires innées dynamiques des cellules épithéliales bronchiques humaines à une infection à coronavirus associée au syndrome respiratoire aigu sévère. PLoS ONE 5 , e8729 (2010).
- Qiu, X. et coll. Réversion de la maladie à virus Ebola avancée chez les primates non humains avec ZMapp. Nature 514 , 47-53 (2014).
- Sui, J. et coll. Élargissement de l'activité de neutralisation pour bloquer directement une voie d'évolution du SRAS-coronavirus dominante dirigée par les anticorps. PLoS Pathog. 4 , e1000197 (2008).
- Sui, J. et coll. Effets des anticorps anti-domaine de liaison au récepteur de protéine de pointe humaine sur l'évasion et l'aptitude de neutralisation du coronavirus du syndrome respiratoire aigu sévère. J. Virol. 88 , 13769-13780 (2014).
- Rockx, B. et coll. Échapper aux effets de neutralisation des anticorps monoclonaux humains in vitro et in vivo fitness du coronavirus du syndrome respiratoire aigu sévère. J. Infect. Dis. 201 , 946–955 (2010).
- Spruth, M. et coll. Un vaccin candidat contre le coronavirus SRAS à virus entier double inactivé stimule les réponses anticorps neutralisantes et protectrices. Vaccine 24 , 652–661 (2006).
- Bolles, M. et coll. Un vaccin contre le coronavirus du syndrome respiratoire aigu sévère à double inactivation fournit une protection incomplète chez la souris et induit une réponse pulmonaire pro-inflammatoire éosinophile accrue lors de la provocation. J. Virol. 85 , 12201-12215 (2011).
- Siegrist, C.-A. in Vaccines 6th edn. (eds. Plotkin, SA, Orenstein, WA & Offit, PA) 14–32 (WB Saunders, 2013).
- Deming, D. et coll. Efficacité du vaccin chez des souris sénescentes soumises à un test de dépistage du SRAS-CoV recombinant portant des variantes épidémiques et zoonotiques. PLoS Med. 3 , e525 (2006).
- Graham, RL, Donaldson, EF & Baric, RS Une décennie après le SRAS: stratégies de contrôle des coronavirus émergents. Nat. Rev. Microbiol. 11 , 836–848 (2013).
- Graham, RL & Baric, RS Recombination, les réservoirs et le pic modulaire: mécanismes de transmission interspécifique de coronavirus. J. Virol. 84 , 3134–3146 (2010).
- Agnihothram, S. et al. Un modèle de souris pour le sous-groupe 2c de bétacoronavirus utilisant un variant HKU5 de la souche de coronavirus de chauve-souris. MBio 5 , e00047-14 (2014).
- Relman, DA Métagénomique, diagnostic des maladies infectieuses et enquêtes sur les éclosions: séquence d'abord, poser des questions plus tard? Confiture. Med. Assoc. 309 , 1531-1532 (2013).
- Kaiser, J. Moratorium sur les études de virologie à risque laisse le travail dans 14 institutions dans l'incertitude. ScienceInsider http://news.sciencemag.org/biology/2014/11/moratorium-risky-virology-studies-leaves-work-14-institutions-limbo (2014).
- Frieman, M. et coll. Déterminants moléculaires de la pathogenèse et de la virulence du coronavirus du syndrome respiratoire aigu sévère dans des modèles murins jeunes et âgés de maladie humaine. J. Virol. 86 , 884–897 (2012).
- Ren, W. et coll. Différence d'utilisation des récepteurs entre le coronavirus du syndrome respiratoire aigu sévère (SRAS) et le coronavirus de type SRAS d'origine chauve-souris. J. Virol. 82 , 1899-1907 (2008).
- Sims, AC et coll. La libération du bloc d'importation nucléaire du coronavirus du syndrome respiratoire aigu sévère améliore la transcription de l'hôte dans les cellules pulmonaires humaines. J. Virol. 87 , 3885–3902 (2013).
- Fulcher, ML, Gabriel, S., Burns, KA, Yankaskas, JR & Randell, SH Cultures de cellules épithéliales des voies aériennes humaines bien différenciées. Méthodes Mol. Med. 107 , 183–206 (2005).
- Roberts, A. et coll. Un coronavirus SRAS adapté à la souris entraîne des maladies et la mortalité chez les souris BALB / c. PLoS Pathog. 3 , e5.
Par Gordon Duff, rédacteur en chef
Veteran Today est bloqué par Facebook, il faut copié/collé ce que je fais quand je veux partager un texte de La Cause du Peuple lui aussi bloqué par Fesse de Bouc ...
RépondreSupprimerNous sommes censurés par tous les outils/Médias sionistes qui détestent la vérité et les opinions contraires, et cela depuis longtemps. Mais nous ne jetterons pas l'éponge.
Supprimerseule la sainte messe en latin fera reculer Satan et les forces mauvaises qui règnent dans le monde pour la perte des âmes ( et aussi des corps) www.torah-injil-jesus.com
RépondreSupprimer